Fat supplementation of dairy cattle diets is a common practice to increase dietary energy and to thereby enhance milk and milk fat yields.
However, the ability of dietary fat supplementation to modulate digestion, metabolism and performance can be influenced by the type of fatty acid (chain length and degree of saturation).
Our understanding of fat supplements is further complicated by the level of fatty acid enrichment, type of protection, physical form and size, degree of esterification, and magnitude of digestibility.
To help unravel this complexity, the McFadden research group uses mass spectrometry-based lipidomics to biochemically map the bovine lipidome (over 1,400 complex lipids), resulting in the discovery of unique metabolic effects attributed to specific fatty acids. The current focus is on the role of ceramide during fat supplementation.
The importance of ceramide
Ceramide is a type of sphingolipid found in tissues and circulation that is formed by saturated fatty acids, including palmitic acid and the nonessential amino acid serine (Figure 1).
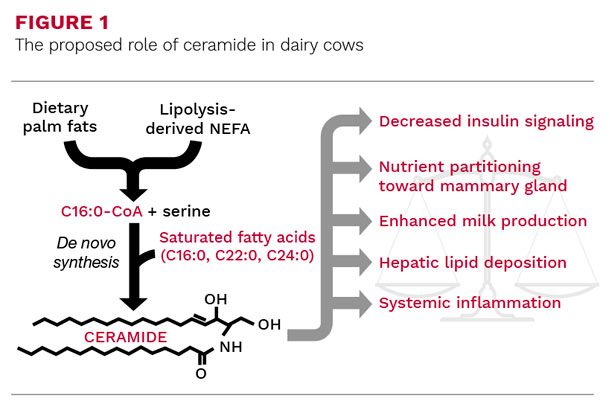
Similar to the diversity of fatty acids, ceramides vary in molecular structure and function. Our understanding of ceramide is primarily attributed to its role as an inducer of insulin resistance, fatty liver, and inflammation in diabetic non-ruminants that consume a caloric-rich diet enriched with palmitic acid.
Albeit, these detrimental effects are highly-dependent on the degree of obesity, and whether ceramide accumulation becomes severe enough to develop into a homeostatic imbalance. This needs emphasis because in the absence of a metabolic disease, ceramide may also serve as an intrinsic control to partition nutrients towards fetal development and lactation.
The potential ability of ceramide to decrease insulin sensitivity and enhance lactation performance in dairy cows are supported by several recent observations.
- First, circulating, liver, and skeletal muscle ceramide concentrations increase in cows transitioning from pregnancy to lactation, but gradually decline beyond peak milk production. Interestingly, increases in circulating ceramide levels develop with adipose tissue breakdown, the systemic accumulation of nonesterifed fatty acids (NEFA) and suppressed insulin concentrations.
- Second, ceramide blocks insulin-stimulated glucose uptake in bovine adipocytes (fat cells) cultured with surplus energy.
- Third, increasing circulating NEFA by intravenous triglyceride infusion or feed restriction increases ceramide and insulin intolerance.
- Fourth, circulating ceramide is consistently positively related to milk yield in dairy cows.
Thus, ceramide has emerged as a novel biomarker for nutrient partitioning in dairy cattle. With this in mind, how might nutritionists modify dietary fatty acid supply to 1) decrease ceramide synthesis in high-risk early lactation cows (cows with high body condition at calving) to potentially enhance insulin action, and lower circulating NEFA and liver lipid deposition or 2) increase ceramide synthesis to spare glucose for the mammary synthesis of milk in mid- or late-lactation dairy cows during positive energy balance?
To explore these questions, investigative effort has focused on the metabolic effects of feeding highly-enriched palmitic acid supplements.
Palmitic acid supplementation increases ceramide synthesis
Palmitic acid is the preferred substrate of serine palmitoyltransferase within the de novo ceramide synthesis pathway (Figure 1). In addition, a second fatty acid is required by ceramide synthase, which is most often saturated, and either palmitic, behenic (C22:0), or lignoceric (C24:0) acid.
Relative to a non-added fat control, feeding early and mid-lactation Holstein cows “high-palm” fats increases ceramide concentrations in blood and liver. Moreover, the ability of palmitic acid to increase circulating ceramide in mid-lactation cows remains superior compared to stearic acid (C18:0) or medium-chain (C8:0 and C10:0) triglyceride supplementation.
The increase in plasma ceramide levels due to palmitic acid supplementation appears to involve hepatic ceramide synthesis and the subsequent secretion of ceramide within very low density proteins. Indeed, palmitic acid feeding increases intracellular ceramide concentrations in bovine primary hepatocytes (liver cells) and circulating low-density lipoprotein ceramide concentrations increase in unison with hepatic NEFA uptake.
Current data suggests palmitic acid feeding promotes nutrient partitioning via mechanisms involving ceramide. In support, circulating NEFA levels remain elevated following a glucose challenge in mid-lactation cows fed palmitic acid when compared with soyhull replacement.
Circulating ceramide is positively related to the postpartum loss in adiposity and the adipose tissue activation of hormone-sensitive lipase. Of potential concern, systemic ceramide concentrations are related to measures of inflammation, including adipose tissue macrophage trafficking and hepatic lipid accumulation during early lactation. However, plasma ceramide concentrations are consistently positively correlated with milk yield in lactating dairy cows that are clinically healthy.
Collectively, these data suggest that a balance between health and milk production must be considered when feeding palmitic acid in early lactation cows at risk of developing metabolic disease. In herds challenged by poor postpartum health and reproductive performance, feeding highly-enriched palmitic acid supplements may be best reserved for beyond peak milk production. Such an approach holds the potential to enhance nutrient partitioning to favor milk synthesis without jeopardizing health.
Current research is focused on the effects of dietary unsaturated fatty acid supplementation on ceramide synthesis. Albeit, our understanding and applications are complicated by their extensive rumen biohydrogenation. Nonetheless, early observations in lambs demonstrate that unprotected palmitoleic (C16:1) acid lowers ceramide supply. This is potentially of related importance because palmitoleic acid enhances insulin sensitivity in obese sheep.
Because dairy cows fed diets containing polyunsaturated fatty acids (PUFA) have reduced pregnancy losses, and dietary PUFA decreases ceramide and increases insulin action in non-ruminants, we hypothesize that PUFA lowers ceramide levels and enhances insulin sensitivity in dairy cows. This possibility is currently being considered in lactating dairy cows at the Cornell University Dairy Research Center in Hartford, New York.
Take-home message
Collectively, ceramide has emerged as a novel biomarker for insulin antagonism in dairy cattle and a target for the development of nutritional strategies aimed at modulating nutrient partitioning. Although studies have repeatedly demonstrated the ability of palm fat supplementation to induce ceramide synthesis in dairy cows, ongoing work aims to delineate the ceramide-related effects of very long-chain saturated fatty acids and PUFA that escape biohydrogenation, and the influence of emulsifiers that may enhance palmitic acid digestibility.
Moreover, we should consider how palmitic acid-induced ceramide accrual is influenced by dietary energy density, co-supplementation with other fatty acids, level of milk production, stage of lactation, and parity. Unraveling these uncertainties has the potential to optimize fatty acid nutrition in dairy cattle.
Joseph W. McFadden is an Assistant Professor of Dairy Cattle Biology, Northeast Agribusiness and Feed Alliance Sesquicentennial Faculty Fellow, in the Department of Animal Science at Cornell University. Email Joseph W. McFadden.
This article appeared in PRO-DAIRY’s The Manager in July 2018. To learn more about Cornell CALS PRO-DAIRY program, visit PRO-DAIRY Cornell CALS.



